
 Empresa
Laboratório de Ensino Zootécnico - UFRGS
Brasil
1
Publicações
78
Seguidores
Empresa
Laboratório de Ensino Zootécnico - UFRGS
Brasil
1
Publicações
78
Seguidores
ESTRATÉGIAS PARA MAXIMIZAR A RETENÇÃO DE NITROGÊNIO E ENERGIA EM SUÍNOS
Com a crescente demanda da sociedade para reduzir a pegada de carbono, os sistemas de produção animal têm se concentrado em aumentar a sustentabilidade da produção de ingredientes para rações e tirar o maior proveito da utilização desses na alimentação animal enquanto promove o descarte correto de resíduos. Assim, mais do que formular rações que reduzam as perdas por excreção de nutrientes, é necessário combinar o uso de diferentes estratégias para maximizar a eficiência ambiental dos programas nutricionais. Estratégias tais como formular rações usando ingredientes locais, usando coprodutos da indústria de alimentos e bioenergia, formulando dietas de baixa proteína ou aumentando o uso de amino ácido (AA) cristalino, e usando variedades mais eficientes reduzindo o uso de fertilizantes (por exemplo, agricultura de precisão) foram propostas por Meul et al. (2012). Entre todas essas estratégias, a agricultura de precisão é uma das mais promissoras para combater as mudanças climáticas. No entanto, ao considerar as mudanças no uso da terra, dietas de baixa proteína usando AA cristalinos parecem ser a estratégia mais eficiente para mitigar a pegada de carbono (Meul et al., 2012). Além disso, em contextos de produção como o da Europa, onde ingredientes para rações são frequentemente importados de países distantes como Brasil e Argentina, reduzir a utilização de farelo de soja diminui significativamente o uso da terra, a pegada de carbono e as emissões de gases de efeito estufa (van der Werf et al., 2005; Garcia-Launay et al., 2014; Garcia-Launay et al., 2018). A redução da utilização do farelo de soja é possível formulando dietas com baixo teor de proteína e incorporando AA cristalino, utilizando alimentação de precisão, ou ambas praticas. No entanto, essas alternativas de alimentação são ambientalmente viáveis apenas se não comprometerem o desempenho do crescimento (Monteiro et al., 2017a; Monteiro et al., 2017b).
Caso teórico: formulando 5 exemplos de dietas
Com o objetivo de criar um caso teórico, sem impacto no desempenho zootécnico, formulamos 5 dietas para estimar as alterações causadas pela redução do teor de proteína bruta (PB) nas dietas. Com base em estudos que abordam o uso de dietas de baixa proteína (Meul et al., 2012; Wang et al., 2018), as dietas foram formuladas usando a matriz nutricional do National Research Council (NRC, 2012) e os coeficientes de digestibilidade do software Evapig® (v. 1.4.0.1; INRA, Saint-Gilles, France) para atender as exigências nutricionais suínos de 25-50 kg de peso corporal alimentados em um programa por fases (Hauschild et al., 2012; Remus et al., 2020a) com base no perfil de proteína ideal (van Milgen and Dourmad, 2015; Remus et al., 2019a). Os ingredientes utilizados para a formulação da dieta basal (Dieta 1) foram milho, trigo, farelo de soja, farelo de canola, óleo vegetal, fontes minerais (pré-mistura micromineral, carbonato de cálcio, fosfato bicálcico hidratado) e fitase. A escolha do ingrediente baseou-se na disponibilidade de produtos locais canadenses, bem como no custo dessas commodities em janeiro de 2021.
Perfil animal teórico: simulando o uso de proteína e energia
O desempenho suíno estimado foi obtido com base em estudos anteriores (Remus et al., 2019a; Remus et al., 2019b; Remus et al., 2020a). Assumindo que em uma fase de crescimento de 28 dias, o animal médio teria um consumo médio diário de ração (CMDR) de 2 kg, ganho de peso médio diário (GMD) de 0,95 kg e uma deposição média de proteína (DP) de 152 g/d. A exigência diária de lisina (Lis; g/d) foi calculada adicionando as exigências de mantença e crescimento (Hauschild et al., 2012). Para um exemplo prático de como usar este modelo, veja o material suplementar provido por Remus et al. (2020a).
Para simular retenção de nutrientes, as exigências de nitrogênio (N) foram assumidas como sendo a quantidade de N retido na DP (DP dividido por 6,25) mais as exigências de mantença proteica (van Milgen et al., 2008). As perdas de energia fecal foram estimadas pela diferença entre a energia bruta e a energia digestível da dieta. As perdas de energia urinária (necessárias para excretar N em excesso e produção de metano) foram estimadas pela diferença entre a energia digestível e metabolizável na dieta. O incremento de calor foi obtido pela diferença entre a energia metabolizável e a líquida da dieta. Esses valores foram multiplicados pelo CMDR para estimar as perdas de energia em Mj / dia. Os valores de excreção de nitrogênio foram obtidos subtraindo a respectiva retenção de nutrientes dos respectivos valores de ingestão de nutrientes.
Resultados do estudo in silico: respostas animais teóricas
Como o milho é pobre em lisina (NRC, 2012), para atender às necessidades de Lis digestível estandardizada ileal, que foi o primeiro AA limitante na dieta, a inclusão de farelo de soja e canola representou 38% da dieta, resultando em 22% de PB (Figura 1). O uso de L-lisina (Figura 1) resultou em uma mudança de 7% na PB na dieta 2 (21% de PB). Devido à combinação de nutrientes fornecidos por nossa matriz nutricional, após a Lis, a treonina tornou-se o próximo AA limitante (Figura 1). A inclusão de soja e canola na dieta 2 diminuiu com o aumento de milho, trigo e adição de AA sintéticos: L-lisina e L-treonina, resultando em uma mudança de 10% na PB na dieta 3 (conteúdo de 20% PB). Como consequência, o triptofano e a metionina se tornaram os próximos AAs limitantes na dieta e, ao suplementar com AA sintético, uma mudança de 17% na dieta 4 (teor de PB de 19%), tornando a valina o próximo AA limitante. Ao suplementar a dieta 5 com L-Lisina, L-Triptofano, L-Treonina, MHA-Metionina e L-valina, uma mudança de 26% na dieta 5 (teor de PB de 16%) em comparação com a dieta 1. É importante enfatizar que a ordem de limitação de AA e a mudança potencial de PB na dieta dependem da matriz nutricional utilizada, que pode variar ao redor do mundo.
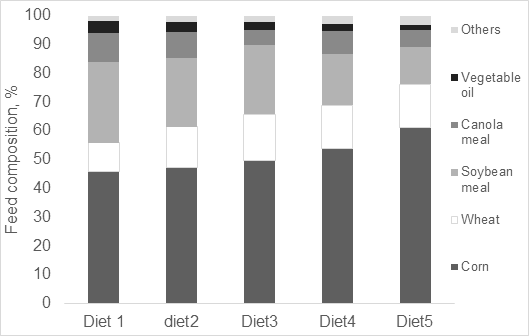
Figura 1. Composição das dietas formuladas para suínos em crescimento (25-50 kg de peso corporal) baseados em ingredientes canadenses, quando os farelos de soja e canola são gradualmente substituídos por milho e aminoácidos sintéticos.
Em termos de mudança de ingredientes, o uso de cinco AA sintéticos permitiu diminuir em 50% o farelo de soja e canola, se comparando a dieta 1 (Figura 1), onde esses alimentos representaram 38% da dieta. Essa mudança na formulação de ração pode ser responsável não apenas por mudanças no uso da terra e, por consequência, diminuição da pegada de carbono (Van der Werf et al., 2005; Garcia-Launay et al., 2014), mas por um aumento da eficiência de utilização de nutrientes também. Wang et al. (2018) descobriram que para cada 10 g/kg de redução da PB na dieta resultará em uma diminuição de 8 a 10% na excreção de N. Para estimar a mudança potencial na excreção de N resultante de mudanças na formulação da dieta no presente estudo, usamos os cálculos do balanço de N conforme apresentado anteriormente. É possível observar (Figura 2) uma mudança progressiva na excreção de N com a redução da PB da dieta. A retenção de nitrogênio pode ser aumentada em 36%, diminuindo de 22 para 16% o teor de PB nas dietas para suínos em crescimento. As mudanças na formulação da dieta também resultaram em uma redução no custo da ração (Figura 2) e a dieta 5 (16% PB) foi 11% mais barata do que a dieta 1, principalmente devido às reduções de farelo de soja e canola na dieta.
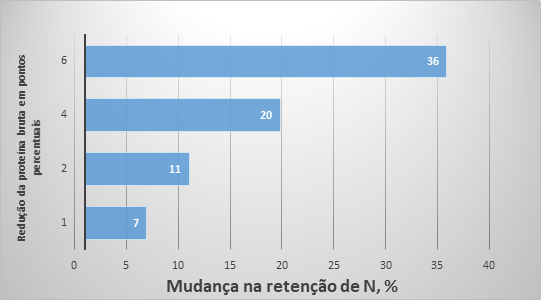
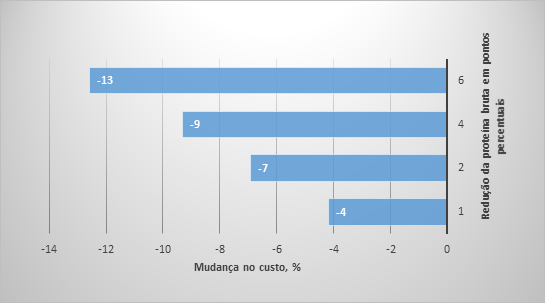
Figura 2. Mudanças na retenção estimada de nitrogênio e nos custos de alimentação como resultado de mudanças no conteúdo de proteína bruta na dieta.
Em termos de eficiência energética de utilização pelo animal (Figura 3), as dietas com baixa proteína também podem apresentar uma vantagem. As perdas fecais de energia são responsáveis pelas perdas de energia após a digestão (Noblet, 2007). E essas perdas podem ser reduzidas em 11% usando dietas com baixo teor de PB (22% vs. 16% de PB). Ao avaliar a diferença da energia digestível para a metabolizável, são contabilizadas as perdas energéticas devido a produção de gás e calor provenientes do intestino posterior (Noblet, 2007). Como o principal gás proveniente desse processo é o metano e responde por apenas 0,4% do N digestível (Noblet, 2007), o principal fator responsável pelas diferenças da energia digestível para a metabolizável está relacionado às perdas urinárias. Portanto, foi para esse componente que as mudanças na formulação da dieta apresentaram a maior diferença no custo energético. Perdas de energia urinária e fermentação intestinal, foram 24% maiores para suínos alimentados com dieta basal (22% PB) do que com dieta baixa em PB (16% PB). Provavelmente porque 31,09 kj/g de N são necessários para excretar o excesso de N na urina (NRC, 1998). O último fator a ser considerado é o incremento calórico da dieta, obtido como diferença entre a energia metabolizável e a líquida. Isso contabiliza os custos relacionados à utilização da energia metabolizável relacionada à ingestão e digestão da ração e o custo da atividade física mínima (van Milgen and Noblet, 2003; Noblet, 2007). O incremento calórico pode ser diminuído em 13% (22% vs. 16% PB) pela diminuição da PB na dieta. Essa mudança no incremento calórico, deve-se principalmente à mudança na proporção do teor de amido e proteína na dieta. Como a glicose é usada de forma mais eficiente do que a proteína como fonte de ATP (van Milgen et al., 2001), ela diminui o incremento calórico. Além disso, os autores sugeriram, de acordo com a literatura (Reeds et al., 1981; Roth et al., 1999), que maior teor de PB nas dietas estimula o turnover de proteína corporal, processo que aumenta o gasto energético.
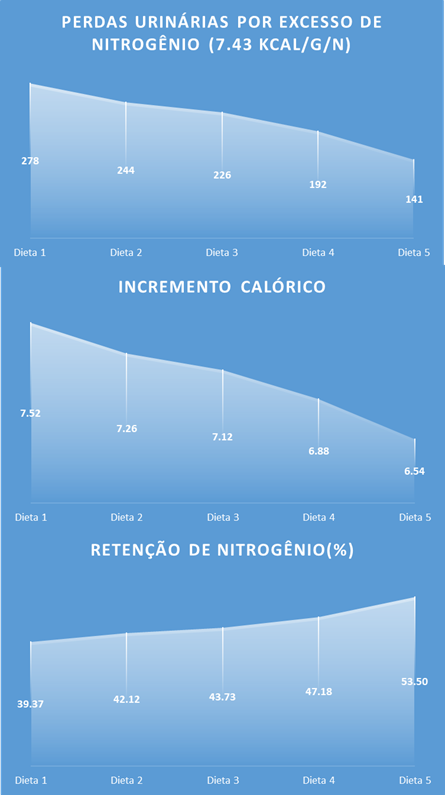
Figura 3. Mudanças no uso da energia da dieta associadas a diminuição de proteína bruta na dieta de suínos em crescimento (25-50 kg).
Alimentação de precisão vs dietas de baixa proteína
Com o objetivo de comparar o consumo de proteína bruta e a eficiência de utilização do N, um exemplo teórico de um sistema de alimentação em 4 fases (Figura 4) foi criado com base nas dietas e cálculos apresentados na Figura 3. Cada fase de crescimento durou 28 dias e que o sistema de alimentação (por fases ou de precisão) e concentração de PB não causam diferença na CMDR, GMD ou DP. A composição da dieta e o desempenho do crescimento para ambos os sistemas foram baseados no trabalho de Andretta et al. (2016). Os cálculos teóricos seguiram a mesma metodologia da seção anterior em dietas de baixa proteína. A ingestão teórica de N em suínos no programa de alimentação individual de precisão (IPF) foi, respectivamente, 20, 40 e 65% menor do que no grupo alimentado com suínos recebendo dietas com baixo, médio e alto PB (Figura 4) nas fases de crescimento e terminação. Essa mudança no consumo de N resultou nos suínos no programa IPF sendo 16, 27 e 40% mais eficientes na retenção de N do que suínos alimentados em grupo recebendo dietas de baixa, média e alta PB, respectivamente (Figura 4). Dietas com baixa proteína parecem ser a alternativa mais próxima para diminuir a excreção de N em sistemas convencionais de alimentação por fase quando comparadas à alimentação de precisão. Ainda assim, a alimentação de precisão oferece benefícios adicionais de redução da excreção de P em grupo alimentado por porcos (Andretta et al., 2014; Andretta et al., 2016). Outro ponto é que se assumirmos que 31,09 kj/g de N são necessários para excretar o excesso de N, as perdas de energia urinária (ignorando a produção de metano) para IPF seriam:
694 kj (perdas de energia urinária) = [(42,8 g N ingestão − 25,1g N necessário para DP + mantença× 31,09 kj] 694 kj perdas energética urinária = 42,8 g N ingestão − 25,1g N necessário para DP + mantença × 31,09 kj
Considerando que a alimentação convencional por fases em grupo seria:
1021 kj (perdas de energia urinária) = [(51,2 g N ingestão − 25,1g N necessário para DP + mantença) × 31,09 kj] 1021 kj perdas de energia urinária = 51,2 g N ingestão − 25,1g N necessário para DP + mantença × 31,09 kj
Resultando que os suínos no programa IPF apresentam 47% menos perdas energéticas do que suínos alimentados em grupo nas 4 fases de crescimento.
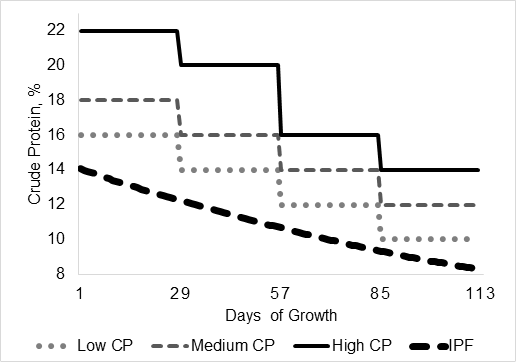
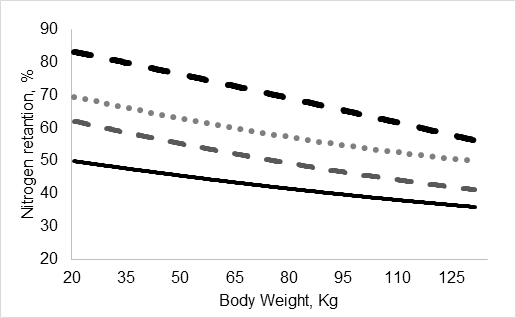
Figura 4. Conteúdo simulado de proteína bruta (PB) na dieta e retenção de nitrogênio (%) em suínos no sistema de nutrição de precisão (IPF) ou no sistema convencional por fase recebendo dietas com conteúdo variável de PB.
Um estudo fatorial in vivo mais recente comparando a IPF com o programa convencional de alimentação em grupo alimentado com dietas reduzidas de PB em ambos os sistemas de alimentação mostrou um ganho aditivo na eficiência de utilização de nutrientes ao usar dietas de baixa proteína dentro do sistema IPF ((Santos et al., 2019; Remus et al., 2020b). Este estudo mostrou que, em geral, nas 2 fases de crescimento estudadas os suínos no programa IPF foram 22 e 29% mais eficientes em reter N e P, respectivamente, do que suínos alimentados em grupo. E a redução da PB nas dietas teve um efeito aditivo no programa IPF aumentando a eficiência de utilização do N em 27%.
Referências bibliográficas
Andretta, I., C. Pomar, J. Rivest, J. Pomar, P. Lovatto, and J. Radünz Neto. 2014. The impact of feeding growing–finishing pigs with daily tailored diets using precision feeding techniques on animal performance, nutrient utilization, and body and carcass composition. Journal of animal science 92(9):3925-3936.
Andretta, I., C. Pomar, J. Rivest, J. Pomar, and J. Radünz. 2016. Precision feeding can significantly reduce lysine intake and nitrogen excretion without compromising the performance of growing pigs. Animal 10(7):1137-1147.
Garcia-Launay, F., L. Dusart, S. Espagnol, S. Laisse-Redoux, D. Gaudré, B. Méda, and A. Wilfart. 2018. Multiobjective formulation is an effective method to reduce environmental impacts of livestock feeds. British Journal of Nutrition 120(11):1298-1309. doi: 10.1017/S0007114518002672
Garcia-Launay, F., H. M. G. van der Werf, T. T. H. Nguyen, L. Le Tutour, and J. Y. Dourmad. 2014. Evaluation of the environmental implications of the incorporation of feed-use amino acids in pig production using Life Cycle Assessment. Livest. Sci. 161:158-175. doi: http://dx.doi.org/10.1016/j.livsci.2013.11.027
Hauschild, L., P. A. Lovatto, J. Pomar, and C. Pomar. 2012. Development of sustainable precision farming systems for swine: Estimating real-time individual amino acid requirements in growing-finishing pigs. Journal of Animal Science 90(7):2255-2263. doi: 10.2527/jas.2011-4252
Meul, M., C. Ginneberge, C. E. Van Middelaar, I. J. M. de Boer, D. Fremaut, and G. Haesaert. 2012. Carbon footprint of five pig diets using three land use change accounting methods. Livestock Science 149(3):215-223. doi: https://doi.org/10.1016/j.livsci.2012.07.012
Noblet, J. 2007. Net energy evaluation of feeds and determination of net energy requirements for pigs. Revista Brasileira de Zootecnia 36:277-284.
NRC, N. R. C.-. 2012. Nutrient Requirements of Swine: Eleventh Revised Edition. 12th ed. The National Academies Press, Washington, DC.
Reeds, P. J., M. F. Fuller, A. Cadenhead, G. E. Lobley, and J. D. McDonald. 1981. Effects of changes in the intakes of protein and non-protein energy on whole-body protein turnover in growing pigs. Br J Nutr 45(3):539-546. doi: 10.1079/bjn19810132
Remus, A., L. Hauschild, E. Corrent, M.-P. Létourneau-Montminy, and C. Pomar. 2019a. Pigs receiving daily tailored diets using precision-feeding techniques have different threonine requirements than pigs fed in conventional phase-feeding systems. Journal of Animal Science and Biotechnology 10(1):16. (journal article) doi: 10.1186/s40104-019-0328-7
Remus, A., L. Hauschild, and C. Pomar. 2020a. Simulated amino acid requirements of growing pigs differ between current factorial methods. animal 14(4):725-730. doi: 10.1017/S1751731119002660
Remus, A., C. Pomar, D. Perondi, J. P. Gobi, W. C. da Silva, L. J. de Souza, and L. Hauschild. 2019b. Response to dietary methionine supply of growing pigs fed daily tailored diets or fed according to a conventional phase feeding system. Livestock Science 222:7-13. doi: https://doi.org/10.1016/j.livsci.2019.02.006
Remus, A., L. S. Santos, C. Pomar, and L. Hauschild. 2020b. Individual Precision Feeding and lowering Crude Protein might help to decrease environmental burden 71th Annual Meeting of the European Federation of Animal Science (EAAP). Wageningen Publishers, Porto Virtual Meeting.
Roth, F., G. Gotterbarm, W. Windisch, and M. Kirchgessner. 1999. Influence of dietary level of dispensable amino acids on nitrogen balance and whole-body protein turnover in growing pigs. Journal of Animal Physiology and Animal Nutrition (Germany)
Santos, L. S., D. Perondi, A. Veira, J. Gobi, and C. Pomar. 2019. PSVII-6 Precision feeding and reduced crude protein on nitrogen efficiency of pigs raised under tropical conditions. Journal of Animal Science 97(Suppl 3):361-361. doi: 10.1093/jas/skz258.721
van der Werf, H. M. G., J. Petit, and J. Sanders. 2005. The environmental impacts of the production of concentrated feed: the case of pig feed in Bretagne. Agric. Sys. 83(2):153-177. doi: https://doi.org/10.1016/j.agsy.2004.03.005
van Milgen, J., and J.-Y. Dourmad. 2015. Concept and application of ideal protein for pigs. Journal of animal science and biotechnology 6(1):1.
van Milgen, J., and J. Noblet. 2003. Partitioning of energy intake to heat, protein, and fat in growing pigs. Journal of Animal Science 81(14_suppl_2):E86-E93. doi: 10.2527/2003.8114_suppl_2E86x
van Milgen, J., A. Valancogne, S. Dubois, J.-Y. Dourmad, B. Sève, and J. Noblet. 2008. InraPorc: A model and decision support tool for the nutrition of growing pigs. Animal Feed Science and Technology 143(1):387-405.
Wang, Y., J. Zhou, G. Wang, S. Cai, X. Zeng, and S. Qiao. 2018. Advances in low-protein diets for swine. Journal of Animal Science and Biotechnology 9(1):60. doi: 10.1186/s40104-018-0276-7
Contato:
Entre em contato conosco através do formulário