

A pele representa um sistema coordenado em que a barreira física, composta pelo epitélio, as células imunes residentes, o microambiente local e o seu microbioma, trabalham juntos para manter a homeostase do corpo.
A pele é composta por três camadas principais e por anexos que incluem folículos pilosos e glândulas sebáceas e sudoríparas (figura 1):

- A epiderme (20-26 μm de espessura), primeira barreira externa, onde a descamação contínua de queratinócitos, a dessecação e o pH baixo, o sebo, os ácidos graxos e as bactérias comensais (até 1012 microorganismos/m2), garantem a sua integridade e dificultam a penetração de agentes patogênicos;
- A derme composta por alguns fibroblastos, fibras de colágeno e elastina, contém sangue, vasos linfáticos e células do sistema imune inato e adquirido;
- A hipoderme, composta por uma camada de gordura, com alguns vasos sanguíneos e linfáticos;
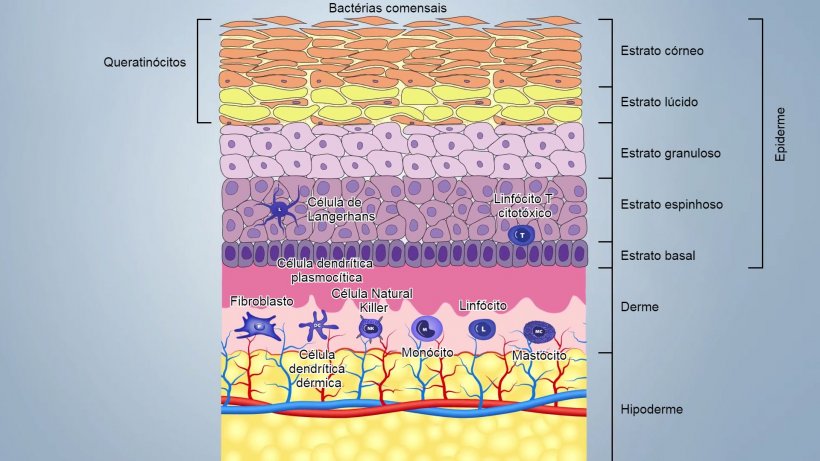
Estas camadas estão estreitamente unidas e formam uma barreira que varia de 50 a 120 μm, dependendo da localização no corpo: nas virilhas e axilas são mais finas que nas outras partes do corpo.
Componentes imunológicos da epiderme
Os queratinócitos são as primeiras células de defensa que expressam uma ampla gama de PRR (Pathogen Recognition Receptors) como TLR (TLR-1, TLR-2, TLR-4, TLR-5 e TLR-6) e lectinas de tipo c. Também segregam uma ampla gama de citocinas pró-inflamatórias e peptídeos antimicrobianos (AMP). De fato, os queratinócitos podem reconhecer PAMP (Pathogen Molecular Pattern) mediante PPR e desencadear inflamação, segregando interleucinas (IL-1), IFN, citocinas, fatores de crescimento, quimiocinas, defensinas. Também podem expressar MHCII e apresentar antígenos.
O antígeno preso na superfície é capturado e internalizado pelas células de Langerhans (CL), um subconjunto especial de células dendríticas epidérmicas localizadas entre os queratinócitos da camada basal e o estrato espinhoso. As CL processam o antígeno e apresentam-no às células T auxiliares próximas. Na epiderme suína, as CL têm uma morfologia típica de células dendríticas (CD) e expressam moléculas de MHCII e moléculas de superfície como CD1, CD172 e CD207.
As CL epidérmicas também são necessárias para manter uma resposta citotóxica eficiente e para a diferenciação de Th17 desencadeada pela secreção de IL-6, IL-1β e IL-23. No suíno, 50-70% das células dendríticas da pele são CL.
A epiderme suína também contém um pequeno número de células γδT, a maioria das quais são células T de memória citotóxica a longo prazo, residentes na pele. Algumas destas células, quando ativadas, podem matar diretamente os queratinócitos infectados e recrutar outras células.
As imunoglobulinas podem atravessar a pele através de transudação e as IgG1, IgG2 e IgA também podem ser sintetizadas localmente.
Vídeo 1. Resposta imune da pele.
Componentes imunológicos da derme
As células dendríticas CD1+ representam as células imunes dominantes e mais importantes da derme. Um sub-conjunto destas células (células dendríticas dérmicas) pode apresentar o antígeno e induzir a resposta primária das células T no gânglio linfático, enquanto que outro sub-conjunto (células dendríticas plasmocitoides) especializa-se na produção de uma grande quantidade de interferón tipo I em resposta ao reconhecimento de ácidos nucleicos virais.
A derme contém uma grande quantidade de linfócitos que está estimada como sendo superior ao número de células na circulação sanguínea. As células T estão dirigidas à derme pelo antígeno leucocitário cutâneo (CLA- cutaneous leukocyte antigen). Os principais sub-conjuntos de células T são CD4+, Th1, Th2 e Th17.
Foi identificado um sub-conjunto de células T circulantes, que são dirigidas à pele e produzem IL-22, sendo capazes de manter a função de barreira em superfícies expostas do corpo.
Os macrófagos (CD14+) são raros na derme, mas os monócitos, que se diferenciam rapidamente em macrófagos, são eficazmente recrutados depois de se produzir uma lesão ou a inflamação.
Na derme também há células natural killers (NK) que eliminam células infectadas por vírus e células cancerosas com uma citotoxicidade não restringida por MHC (Major histocompatibility complex). As NK são ativadas por interferons ou outras citocinas liberadas por macrófagos ou queratinócitos. As células NK expressam receptores inibidores (KIR- killer cell immunoglobuline-like receptor) que reconhecem o MHC-I e evitam ataques indesejados sobre o próprio organismo. Podem matar as células-alvo através da via perforin-granzima.
Os mastócitos (MC) também são células importantes implicadas na inflamação em resposta aos agentes patogênicos. Os MC agem como células sentinelas capazes de reconhecer sinais de perigo através de TLR e liberarem rapidamente mediadores da inflamação tais como a histamina, leucotrienos, prostaglandina, citocinas pro-inflamatórias e imunomoduladoras (IL-1, IL-3, Il-5, IL-6, GM -CSF, IL-4, IL-10, IL-13) e muitas quimiocinas. Esta atividade pode ser induzida por diferentes estímulos inflamatórios dependentes de IgE e não dependentes de IgE. Os mastócitos também podem funcionar como células apresentadoras de antígeno MHC II restringidas e expressam moléculas específicas (B7-1, B7-2, CD40) para a ativação dos linfócitos T.
Os produtos procedentes de parasitas e bactérias podem ser internalizados e processados pelos mastócitos e depois apresentados aos linfócitos T.
Mais especificamente, durante as reações de hipersensibilidade de tipo I (alergia) os mastócitos participam na resposta inflamatória, liberando mediadores quando as IgE específicas do alérgeno unidas a receptores de imunoglobulina E de alta afinidade (FcεRI) na sua superfície celular, reagem a uma segunda exposição ao mesmo alérgeno.
